 基因君官网
基因君官网
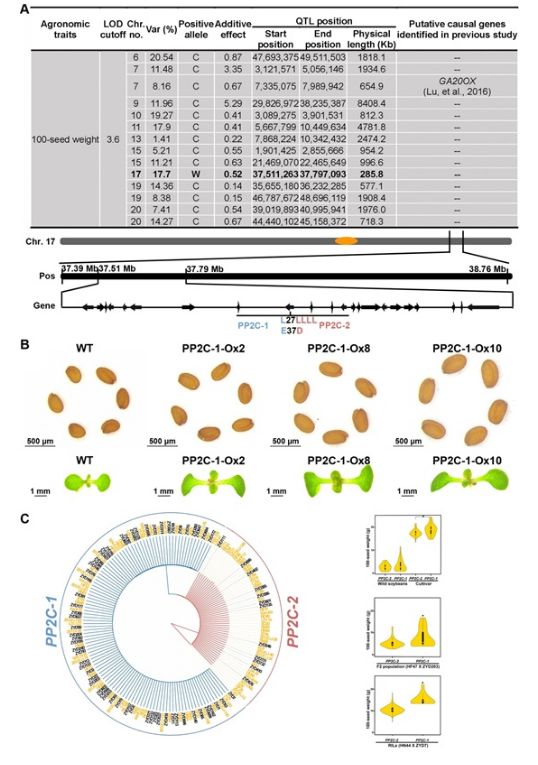
图:大豆百粒重基因鉴定。(A): 利用重测序群体定位的百粒重 QTL 位点。C:表明优势位点来源于栽培大豆 HN44;W:表明优势位点来源于野生大豆 ZYD7。PP2C- 1 在第 27 位氨基酸是 L(亮氨酸),37 位是 E(谷氨酸)。而 PP2C- 2 在同样位置分别有 4 个 L,一个 D(天冬氨酸)。(B): PP2C- 1 在模式植物拟南芥中过表达可使籽粒及子叶变大。WT 为对照,其它为转基因株系。(C): 不同大豆资源中 PP2C 变异分析。左图:近 40% 的大豆资源中没有 PP2C- 1 优异等位变异;黑色名称材料为野生大豆,黄色名称材料为栽培大豆。右上图:野生和栽培大豆中基因型与百粒重的关系;右中图:野生大豆 ZYD203 与栽培大豆 HF47 的 F2 群体中基因型与百粒重相关性比较;右下图:野生大豆 ZYD7 与栽培大豆 HN44 的重组自交系群体中基因型与百粒重的关系。
大豆是我国重要的粮食作物和经济作物,是植物蛋白和油分的重要来源。百粒重是大豆产量的重要构成因子,因此是大豆育种的重要目标性状。由于栽培大豆品种遗传基础狭窄,在育种过程中某些栽培大豆品种中优异等位的丢失,阻碍了大豆百粒重和产量的进一步增加。近年来研究人员对大豆百粒重遗传位点的研究较多,目前 SoyBase 数据库已经收录了定位的百粒重 QTL 位点 94 个。尽管大豆百粒重种质资源筛选和遗传研究工作开始很早,但有关大豆百粒重基因的克隆和种子形成的分子机制研究还较少。
中国科学院遗传与发育生物学研究所研究员张劲松和陈受宜领导的研究团队与黑龙江省农科院大豆研究所研究员满为群和耕作栽培研究所研究员来永才等团队合作,通过分析衍生于野生大豆 ZYD7 和栽培大豆 HN44 的 1036 份重组自交系材料,鉴定了一个种子百粒重增加的株系 R245。随后对 198 份自交系材料和 2 个亲本进行了全基因组重测序,获得了高质量的 SNPs 并作为分子标记,构建了大豆遗传图谱并利用多年多点的数据定位了调控种子百粒重的 14 个 QTL 位点和控制油分含量的 3 个 QTL 位点。在调控百粒重的位点中,13 个位点的优势基因来源于栽培大豆 HN44,1 个位点的优势基因来源于野生大豆 ZYD7(图 1A)。通过对高百粒重的优异品系 R245 进行全基因组重测序及基因分型验证,发现它含有前述的来源于栽培型的 13 个百粒重调控位点及来源于野生型的 1 个百粒重调控位点。来源于野生大豆 ZYD7 的百粒重遗传位点位于 285 kb 区间内,共有 22 个蛋白编码基因,但它们在栽培和野生大豆种子发育期间无显着表达差异。而其中 Glyma17g33690 (PP2C) 和 Glyma17g33790 (EAL) 的核苷酸变异改变了氨基酸序列(图 1A)。转基因分析发现,来源于 ZYD7 的 PP2C- 1 基因显着增加了转基因拟南芥种子的重量,而来源于 HN44 的 PP2C- 2 和 EAL- 2 以及来源于 ZYD7 的 EAL- 1 并不能改变转基因拟南芥种子重量(图 1B)。功能分析发现,PP2C- 1 能与油菜素内酯 BR 信号通路的转录因子如 GmBZR1 等相互作用,通过去磷酸化从而激活这些转录因子,促进下游控制种子大小的基因表达以提高粒重。而 PP2C- 2 则没有上述功能。种质资源分析发现,近 40% 的栽培大豆还没有 PP2C- 1 基因型(图 1C),后续研究可以将 PP2C- 1 基因型通过杂交导入到不含该位点的大豆品种中,从而提高产量潜力,将对于大豆育种具有重要意义。
这项工作于 3 月 28 日在线发表于《分子植物》(Molecular Plant,DOI: 10.1016/j.molp.2017.03.006),该团队的博士生陆翔和熊青为共同第一作者,野生大豆 ZYD7 和栽培大豆 HN44 重组自交系群体由黑龙江省农业科学院大豆研究所的杜维广和满为群团队构建,该研究得到了中科院先导专项和国家转基因专项等项目的大力资助。(生物谷 Bioon.com)